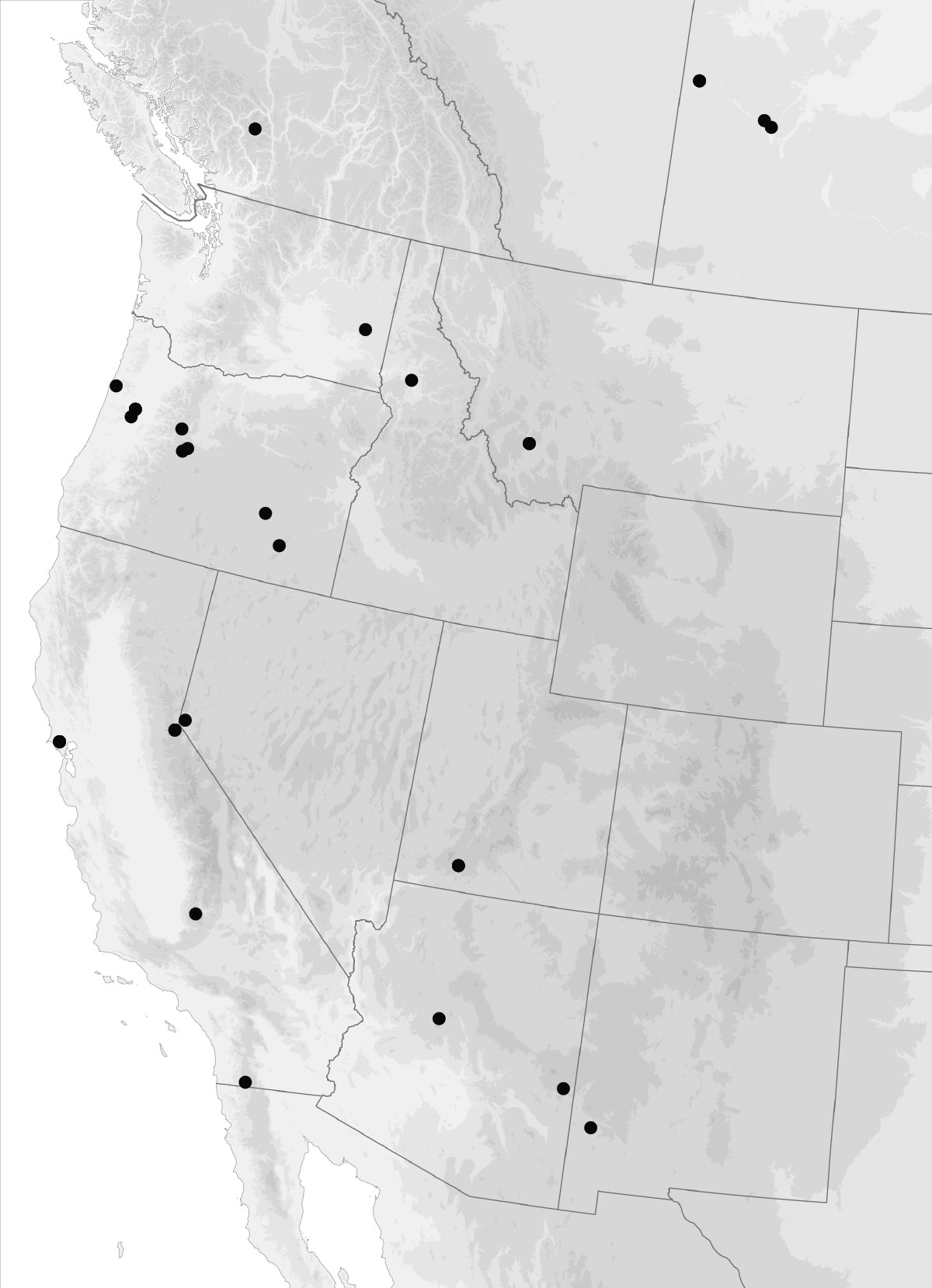
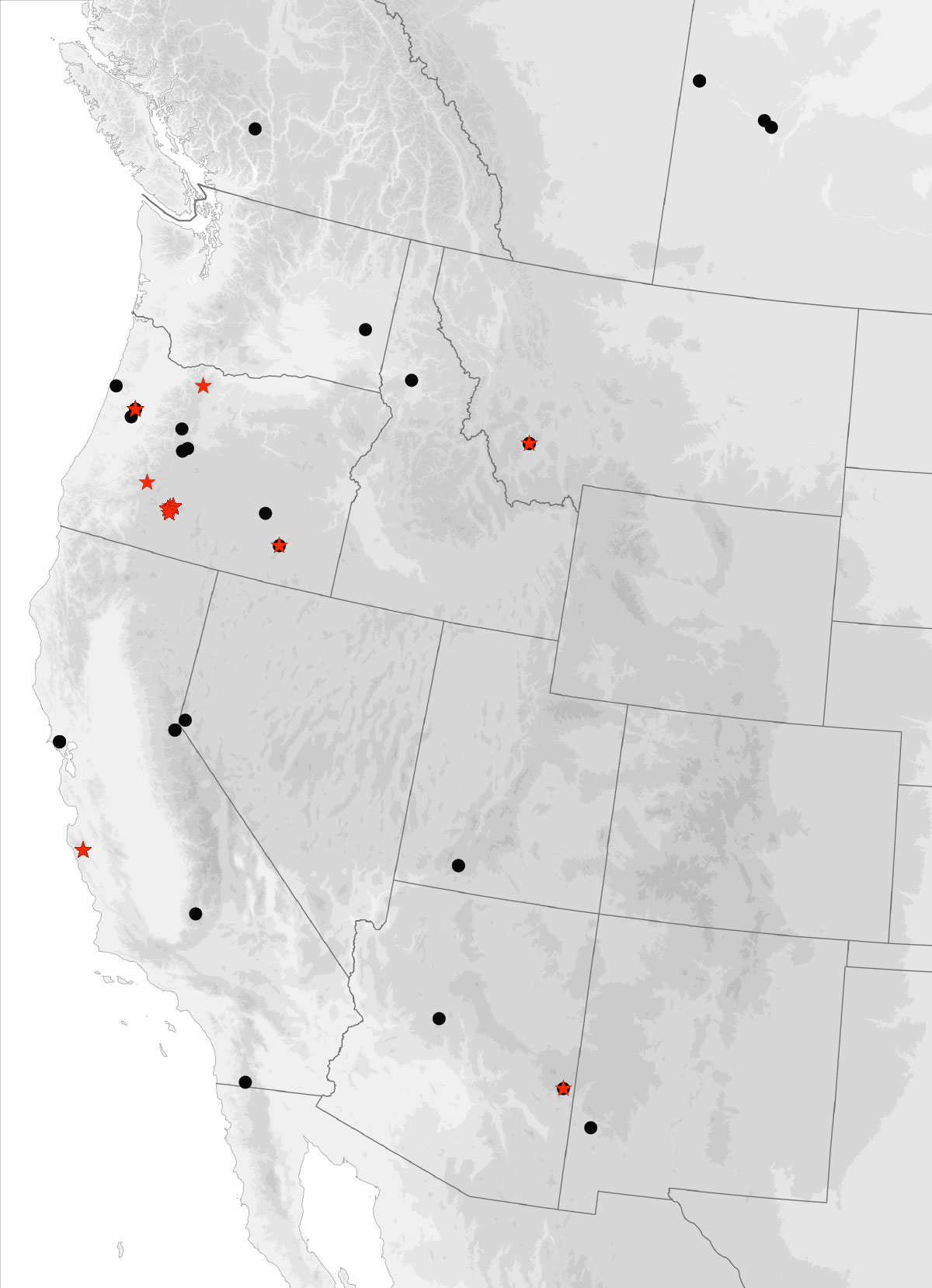
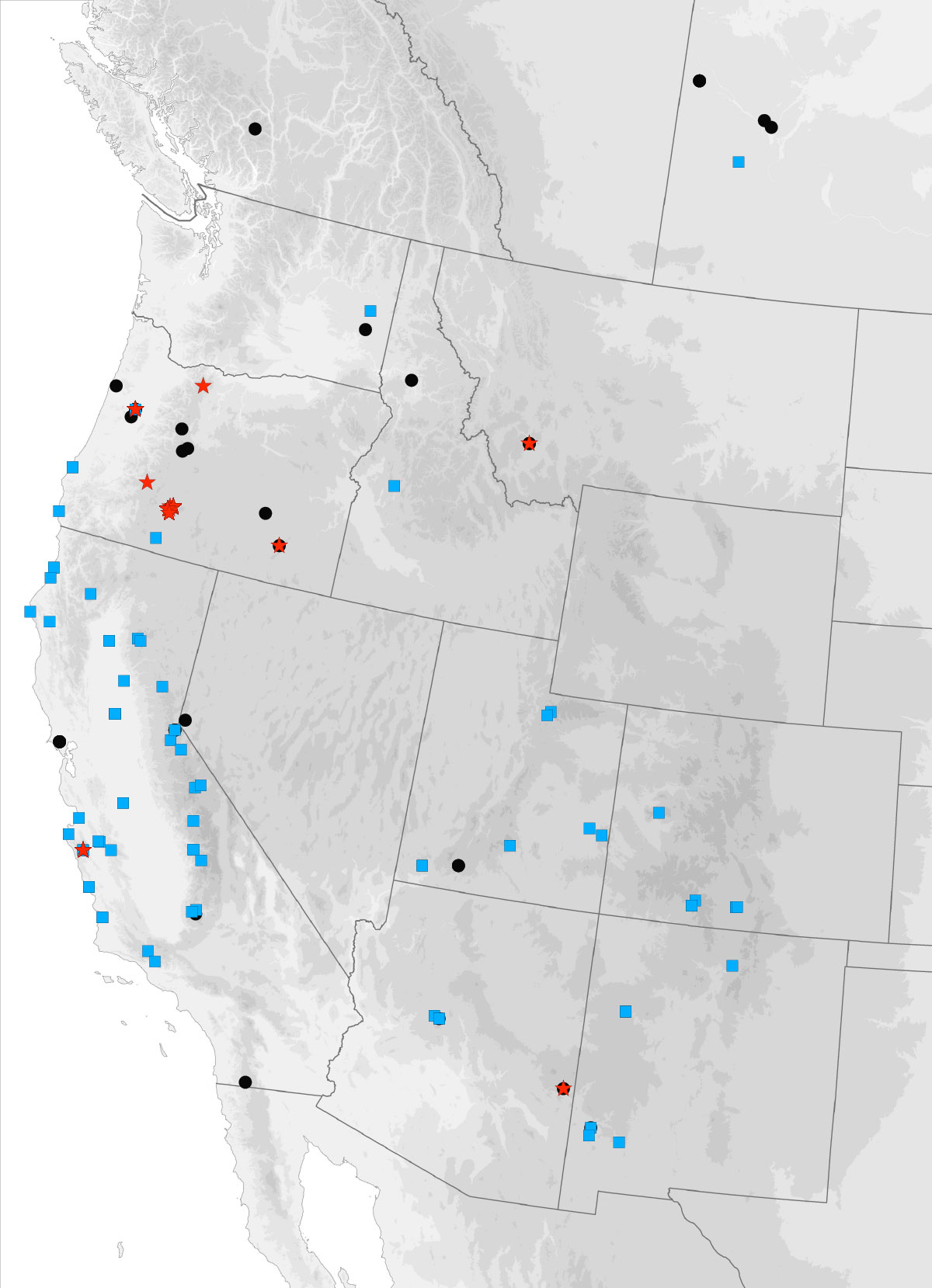
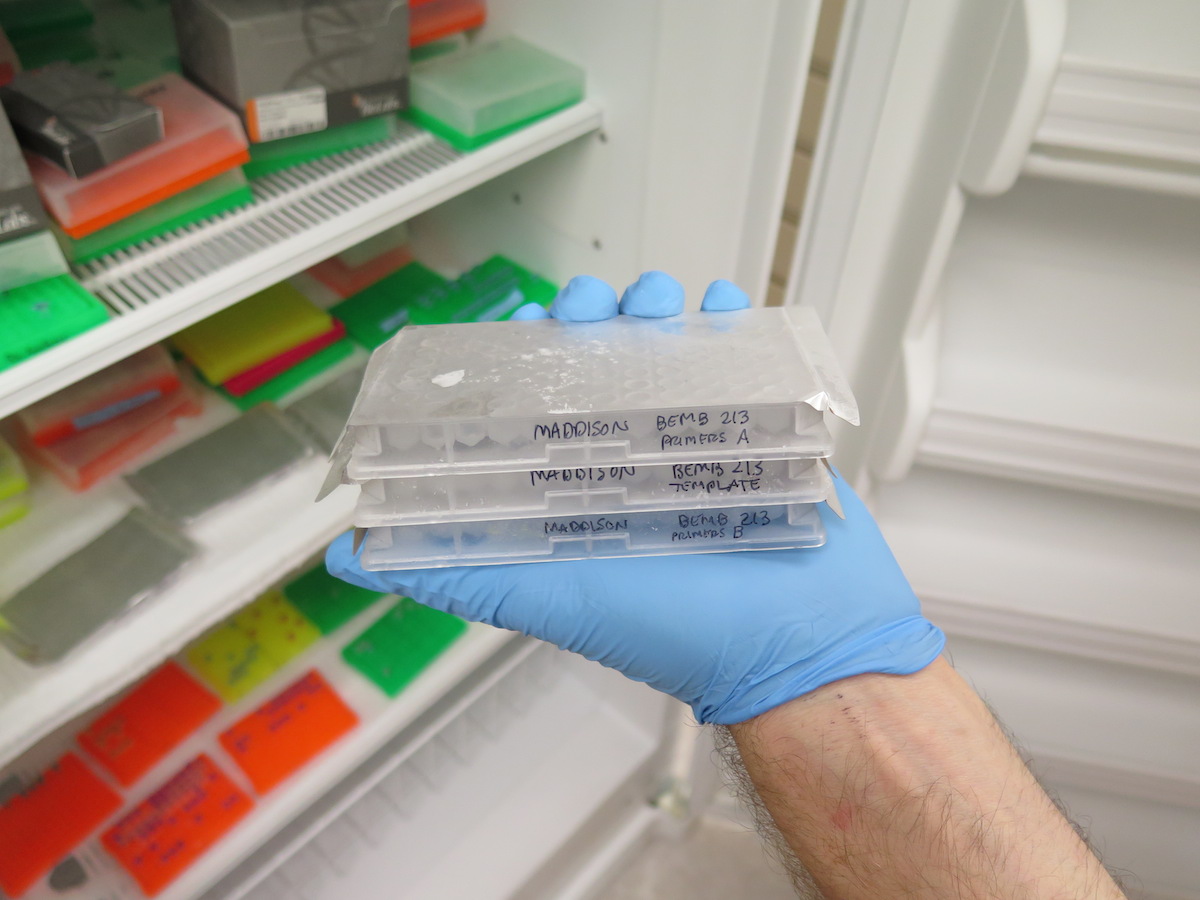
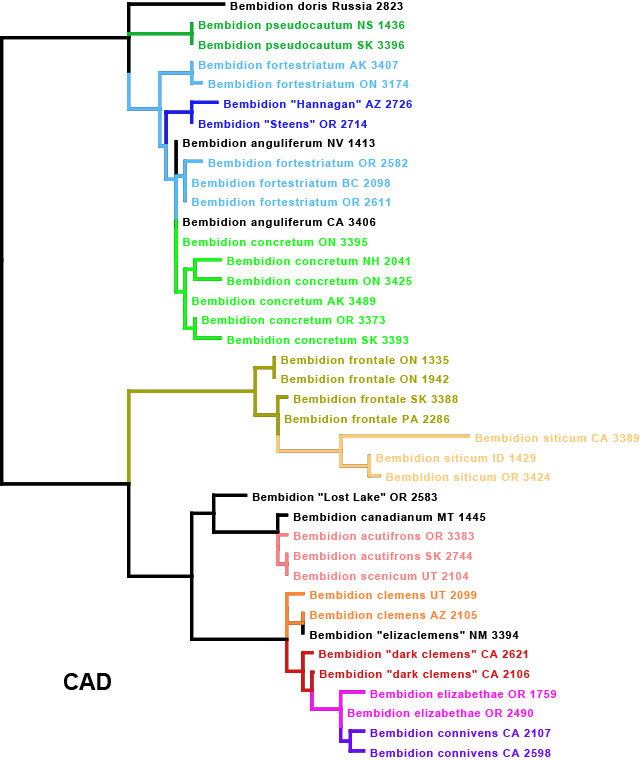
I got the email from FedEx saying that our package of four plates of 384 PCR samples had arrived at the University of Arizona, where they will be sequenced. These plates contain the DNA from multiple genes from over 120 Trepanedoris that we have not sequenced before; the data we get back will tell us a huge amount about Trepanedoris diversity for our Discovering Insect Diversity class.
We send all of our PCR samples to the University of Arizona Genetics Core to be sequenced. They are relatively inexpensive, and do a great job. In particular Stacy Sotak has been terrific, and has helped us over the years with all of my Sanger sequencing needs. In the rare times that there have been problems, Stacy has really worked well with us to help troubleshoot. I worked with Stacy for some of my time at the University of Arizona, and since I moved to Oregon State University she has continued to be a fantastic (if now more distant) help to our research.

I asked Stacy if she could take pictures of the processing of this set of four plates, so that the class could see how the magic happens of going from PCR product to DNA sequences. I’m excited to see some of the equipment used, as I haven’t actually ever seen it myself.
The plates arrived in good shape to the U of A; here they are coming out of the shipping box!

The PCR products then needed cleaning; here are the first two plates (Bemb213 and Bemb214) sitting in the cleaning system:
 After cleaning, the amount of DNA in each of the 96 wells of each plate was then quantified. I looked over the details, and they all seem to have a good amount of DNA in them, which is excellent!
After cleaning, the amount of DNA in each of the 96 wells of each plate was then quantified. I looked over the details, and they all seem to have a good amount of DNA in them, which is excellent!
Update 1: The DNA in the plates have been quantified. Here they are getting ready to be quantified:

And here is the fluorometer that quantifies the DNA:

Update 2 (1 May, about 10:30 am): The sequencing is done! Here is a picture from Stacy showing the machine that does most of the magic, the sequencer itself:

The instrument is open, and on the right you can see six of the eight plates loaded into it.
You might wonder why there are eight plates rather than four – after all, there are only four plates of PCR products. We sequence each sample twice (once in one direction, once in the other direction) so that we can confirm the results. That’s why we had two primer plates for each PCR sample plates. This means we sequence each original plate twice, which translates into eight sequencing plates in total.
Here’s what the computer attached to the sequencer is showing as the sequencing is happening:

When it was all done, Stacy did some data processing, pressed some buttons, and I got the following emails, which always cause my heart to race in excited anticipation, as they are telling me that the sequences are ready to download:
Alas, I will have to wait to download them. I would normally spend today processing the data and doing analyses, and lose myself in the thrill of all of the cool stories the sequences will tell. However, I have promised myself that I will not do this until class on Tuesday – I want the students to be fully part of the process, and see the results the same time I do. For the next four days, I will need to remind myself that patience is a virtue.
[Thanks so much, Stacy, for taking these pictures for the blog – they are much appreciated by the students!]