
In Lindroth’s magnificent 1963 treatment of Bembidion of Canada and Alaska (and the northern contiguous States), he notes the extent of structural variation within species. Some species he notes to be relatively uniform, others more variable. As I delve into bembidiines of North America more thoroughly, I find that just about any species he notes to be rather variable I will find (using more detailed morphological or molecular data) to be multiple species. For example, he starts his description of “Bembidion chalceum” with “An extremely variable species: in size, impression and punctuation of elytral striae, convexity of intervals (including the 9., with seeming variation of its width, as a consequence), &c. ” My 2012 paper on subgenus Pseudoperyphus shows that his “Bembidion chalceum” consists of at least four species, now named Bembidion chalceum, B. rothfelsi, B. bellorum, and B. louisella.
For this reason, I was suspicious of Bembidion kuprianovi, as Lindroth begins the description of that species with the statement: “Varying in size more than perhaps any other species of Bembidion, also in color, form of prothorax (figs. 133a, b), &c.” Some are small and brown (as in the specimen on the left, below), some are large and black (as the specimen on the right), but there seem to be lots of in-betweens.

“Bembidion kuprianovi”
I had become convinced that this was a complex of species in 1981, when I was on a collecting trip to the Yukon with George Ball and Nigel Stork. One day I found a population of B. kuprianovi on damp soil under leaf litter under small willow bushes on the upper bank of a slowly-moving river. All specimens there were the small, pale form of B. kuprianovi.
A few days later, on the gravel shore of a cold, clear, fast river, I found another population of B. kuprianovi, and all of these were of the large, dark form. The strong correlation between body color and form and habitat convinced me right then that there were at least two species involved. Carabid beetles don’t seem to be that ecophenotypically variable the way other organisms such as some plants can be; that is, they seem to develop into similar forms independent of the environment. Thus, it seems unlikely that the different color and sizes are caused by their developing in different environments, and it is more likely that they are genetically different. I never explored this issue further until this last week.
10 days ago I was collecting just east of Moscow, Idaho, along the East Fork of the Potlatch River, with Chris Marshall. Here’s the habitat:

East Fork Potlatch River, Idaho
I found one of the big dark B. kuprianovi close to the water, and a series of pale, small B. kuprianovi up under some willow bushes. I’ve extracted DNA from the big dark one and one of the small, pale ones, but I haven’t sequenced those specimens yet (although I have sequenced other specimens, as I mention below). But I have looked at the male genitalia, as well as those of other large, dark and small, pale ones, and there is an obvious pattern.
In the following series, the top two are the genitalia of two of the large, dark males, and the bottom two are of the small, dark males.

Specimen DNA2101 (large and dark), Alberta

Specimen DNA3300 (large and dark), Idaho

Specimen DNA3082 (small and brown), California
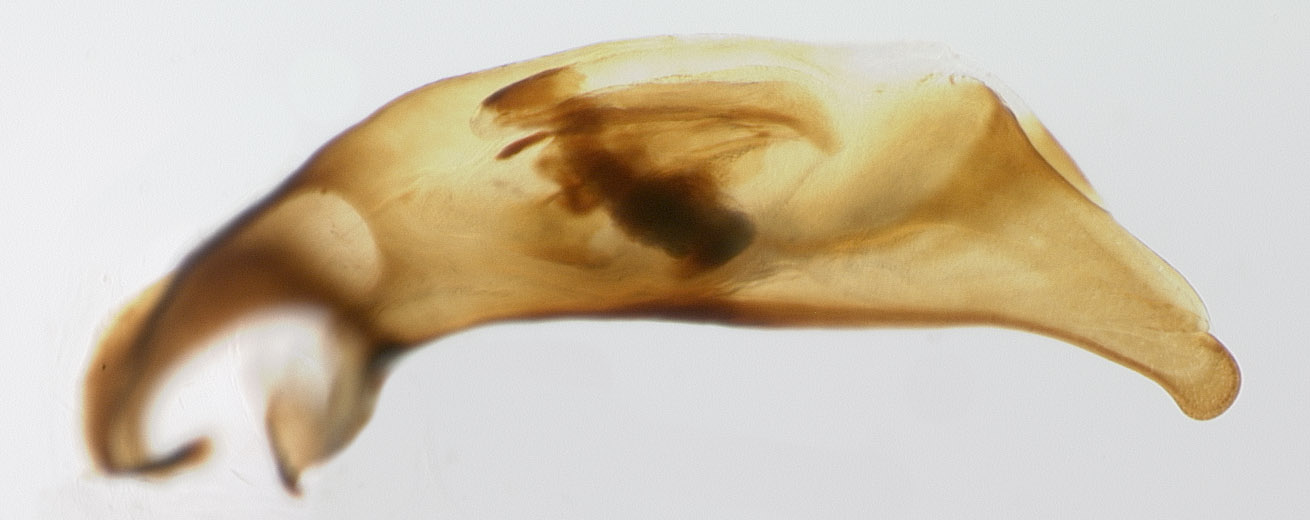
Specimen DNA3299 (small and brown), Idaho
As you can see, the large and dark specimens have longer and thinner aedeagi, with more elongate tips; there are some differences in the internal structures as well. The evidence that there are two different species is thus getting stronger and stronger: the correlation between color, size, habitat, and genitalic structure is quite compelling.
I have sequenced the COI gene for 2 of the small pale specimens, and 2 of the large dark ones. There are no differences between them. I have also sequenced 28S rDNA from those same specimens, and there is a single base difference over the 950 bases sequenced. While the single base difference is not that great, the correlation between that and the other characteristics provides additional confirmation.
Thus, what we now call “Bembidion kuprianovi” is at least two species. One of them will presumably keep the name Bembidion kuprianovi. I suspect the other species already has a name: there are 7 synonyms of Bembidion kuprianovi as it now stands, and one of these names likely applies to the second species. Only examination of the type specimens will reveal which names should be applied to which species.
I should note that the original discovery of the correlation between habitat and body form is an example of the importance of taxonomists going out into the organisms’ habitats and encountering them there. Observing the beetles in nature can allow one to see correlations that provide the key to seeing the complex diversity in just the right way to reveal the pattern of species boundaries.

